ГЕОМЕТРИЧЕСКИЕ МОДЕЛИ В БИОЛОГИИ: КАК И ЧТО МОЖНО МОДЕЛИРОВАТЬ
В.К.БОЖЕНКО*, А.О.ИВАНОВ, А.С.МИЩЕНКО, А.А.ТУЖИЛИН, А . М . ШИШКИН*
Механико-математический факультет МГУ им. М.В. Ломоносова , Россия
*
Российский Научный Центр Рентгенорадиологии МЗ РФ
Оглавление
Цель настоящей статьи – проиллюстрировать возможности геометрического моделирования на примере его использования в биологии. Важность геометрического подхода для наглядного представления скрытых закономерностей, управляюших функционированием живых структур, объясняется не только их сложностью , но и тем, что многие жизненно важные свойства того или иного организма существенно зависят как от его формы в целом, так и от форм составляющих его частей. При этом геометрические особенности проявляют себя на всех уровнях структуры, начиная с макроуровня и заканчивая взаимодействием отдельных молекул. Даже социальные взаимодействия, в которых важную роль играют априорные симпатии, во многом определяются внешним видом контактирующих индивидов, что объясняется геометрической визуализацией свойств, отвечающих за жизнеспособность и адекватность возникающих контактов. В данной работе мы сосредоточим основное внимание на проявлениях геометрии в микромасштабе, а именно, в мире биологических молекул и совокупностей клеток. Дополнительную информацию можно найти в работах [2-6].
Начнем с краткого обсуждения того, как геометрические представления помогают в научных исследованиях. Отметим, что основная задача по сути любого естественнонаучного исследования – догадаться и сформулировать гипотезы, описывающие свойства и взаимосвязи рассматриваемых объектов. Использование визуальных компьютерных моделей позволяет перевести набор формальных определений в наглядные геометрические свойства, за которыми можно наблюдать, динамически меняя параметры. Последнее дает возможность непосредственно увидеть свойства и взаимосвязи, что приводит к формулировке гипотез, далеко не очевидных без компьютерного эксперимента и геометрической визуализации. Конечно же, далеко не все гипотезы превращаются в теоремы. Чтобы проверить справедливость найденных связей, ученые все больше и больше используют компьютерный эксперимент для того, чтобы или построить контрпример, или найти подсказки, которые позволят доказать гипотезу. При этом контрпримеры также являются важным этапом исследования, потому что позволяют более глубоко осознать изучаемые объекты и уточнить формулировки гипотез.
Ниже мы покажем, как можно использовать геометрическое моделирование и визаулизацию на нескольких примерах из биологии: моделирование «химической лаборатории» живой клетки; изучение формы (конформации) биополимеров таких, как ДНК, РНК и белки; исследование формообразования различных тканей в живом организме.
1. Симулирование внутриклеточного взаимодействия
Одной из важнейших задач современной молекулярной биологии является изучение взаимодействия белков и нуклеиновых кислот, регулирующего важные внутриклеточные процессы. Такие процессы обычно моделируются с помощью дискретных динамических систем на графах с весами на ребрах и вершинах, которые пересчитываются по некоторым правилам. Сложность такого моделирования объясняется большим количеством молекул, участвующих во взаимодействии, а также недостатком информации о его природе и законах. Одним из возможных подходов является построение самонастраивающейся многопараметрической модели, в которой бы реализовались хорошо известные глобальные свойства жизнедеятельности клетки такие, как последовательность и продолжительность фаз клеточного цикла и др. При работе с такими моделями особую роль играет наглядность представления поведения весов при динамическом изменении параметров системы.
Нами была разработана программа, позволяющая изучать внутриклеточные взаимодействия. Фактически с помощью этой программы можно строить графы, приписывать ребрам и вершинам векторные веса, визуализировать эти веса цветными столбиками, расположенными рядом с вершинами и ребрами, задавать интерактивно законы пересчета этих весов во времени, наблюдать изменение во времени векторных характеристик графов. Особенностью нашей компьютерной программы является возможность изменения всех ее параметров, включая структуру графа, прямо во время работы динамического модуля, что позволяет эффективно управлять динамикой, добиваться требуемого поведения системы. На рис. 1 показано одно из рабочих окон программы.
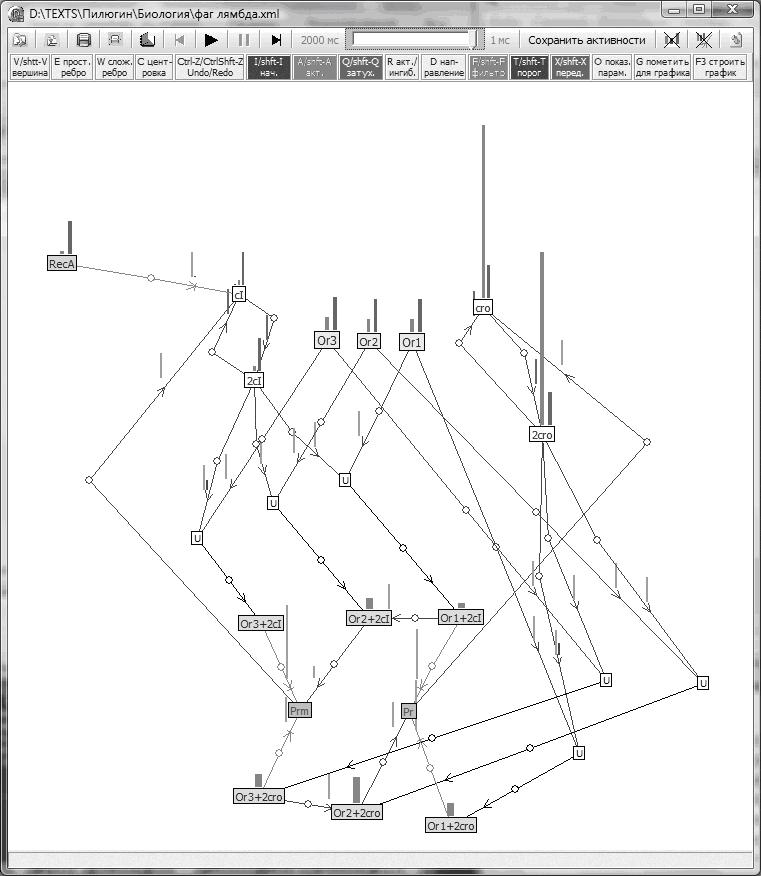
Рис. 1 Моделирование внутриклеточных взаимодействий
2. Геометрия кривых и ее приложение к исследованию конформации полимеров
Глобальная проблема современной биологии – понять, как функционируют живые клетки. Основными объектами живых клеток являются белки и нуклеиновые кислоты. Одно из приложений – исследование пространственной структуры (так называемой конформации) этих биополимеров в терминах их химических формул. Важность изучения конформации объясняется существенным влиянием последней на каталитические свойства. Практическим примером использования математического моделирования конформации макромолекул являются достижения в области фармакологии, основанные на моделировании активных центров белков-мишеней (например, киназы, входящей в комплекс циклина D ) и трехмерной структуры потенциальных ингибиторов. Применение такого подхода позволило получить, спектр перспективных противоопухолевых препаратов – низкомолекулярных ингибиторов пролиферации (см. Annals of oncology . 2007. v .18.- Sup .4.- iv 27).
Непосредственное математическое моделирование, основанное на законах классической или квантовой физики, описывающих взаимодействие атомов, составляющих молекулы, оказывается невозможным благодаря огромному количеству этих атомов. Один из подходов – использование геометрических эвристик, тесно связанных с конформацией. Для исследования таких эвристик требуется обширный компьютерный эксперимент, в частности, визуализация различных геометрических свойств приближений и изучение проявления этих свойств в разных биомолекулах.
Рассмотрим частный случай задачи – изучение конформации белка. Напомним, что основу белков живых клеток составляют аминокислоты, соединенные в линейную цепь. В подавляющем большинстве клеток имеется ровно 20 типов аминокислот, формирующих белки. Пространственная структура белка представляет собой углеродно-азотную линейную цепь, к которой крепятся атомы водорода H , кислорода O , а также аминокислотные остатки R i , характеризующие ту или иную аминокислоту (по одному остатку на кислоту, см. рис. 2).
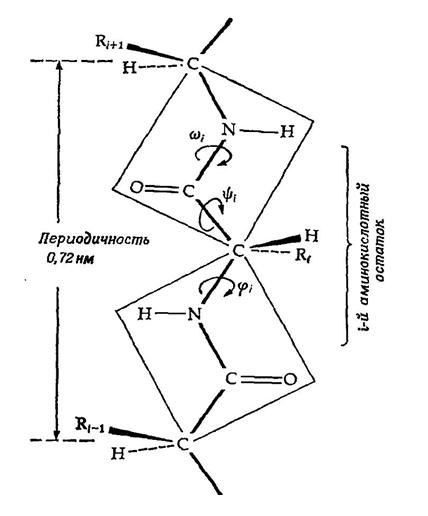
Рис. 2. Фрагмент структуры белка.
Углероды из углеродно-азотной цепи, к которым крепятся аминокислотные остатки, традиционно называются альфа-углеродами.
Соединим последовательные вдоль цепи альфа-углероды прямолинейными отрезками (в масштабах задачи атомы можно считать точками). Полученную ломаную будем называть характеристической . Конформацию белка будем описывать геометрией именно такой ломаной.
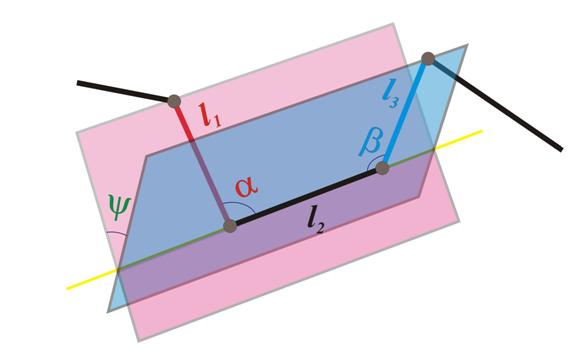
Отметим, что в белках живых клеток расстояние между соседними альфа-углеродами практически одинаково. Тем самым, характеристическая ломаная является равнозвенной, и значит, при известной длине ребра для ее описания достаточно знать углы между последовательными звеньями (углы первого типа), а также между плоскостями (углы второго типа), натянутыми, построенными для каждого некрайнего звена e , так: первая из плоскостей натянута на e и предыдущее звено, а вторая – на e и последующее звено ( см. рис. 3, здесь ? и ? – углы первого типа, а ? – угол второго типа ) . Отметим, что иногда углы первого типа называют валентными , а второго – торсионными .
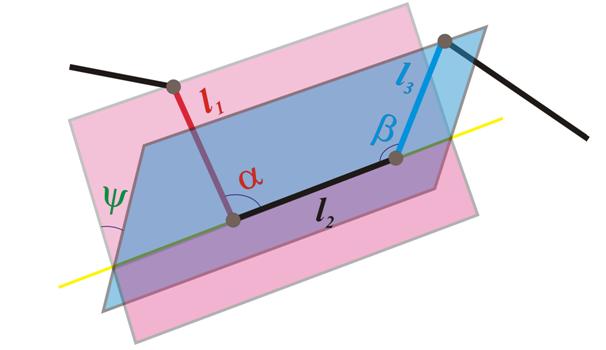
Рис. 3 Углы первого и второго типов
Чтобы лучше понять, как свойства углов связаны с геометрией ломаной, полезно провести аналогию между ломаными и кривыми. Хорошо известно, что бирегулярные пространственные кривые ?(s) определяются однозначно (с точностью до движения) двумя фунциями параметра s – кривизной k(s) и кручением ?(s) . Напомним определение этих функций.
Пусть ?(s) =(x(s),y(s), z ( s )) – натурально параметризованная гладкая пространственная кривая, т.е. функции x(s) , y(s) и z ( s ) непрерывно дифференцируемы нужное число раз, и абсолютная величина скорости ?'(s) =(x'(s),y'(s), z '( s )) движения по кривой равна 1.
Положим v(s) = ?'(s) – скорость (ее величина равна 1), k ( s )=| ?''( s )| – кривизна , n ( s ) = ?''( s )/| ?''( s )| – главная нормаль (мы предполагаем, что кривизна отлична от нуля, т.е. кривая бирегулярна ), b(s) = [v(s),n(s)] – бинормаль (здесь квадратные скобки обозначают векторное произведение). Легко показать, что векторы b'(s) и n(s) пропорциональны. Коэффициент пропорциональности ?(s) между этими векторами, взятый со знаком минус, называется кручением кривой ?(s) . Таким образом, b'(x)= - ?(s)n(s) . Хорошо известен следующий классический результат.
Теорема.
Для каждой положительной гладкой функции f(s) и произвольной гладкой функции g(s) существует пространственная натурально параметризованная кривая ?(s) с кривизной k(s) , равной f(s) , и кручением ?(s) , равным g(s) . Все такие кривые отличаются друг от друга на движение пространства. Плоские кривые однозначно определяются своей кривизной.
Приведем примеры плоских и пространственных кривых с заданными функциями кривизны и кручения (для пространственных кривых):
• если k(s) = 0 , то ?(s) – прямая линия. В случае пространственной прямой кручение не определено;
• если ?(s) = 0 , то ?(s) – плоская кривая;
• если для плоской кривой выполнено k(s) = const > 0 , то она является окружностью. Для пространственной кривой окружность получается при добавлении условия ?(s) = 0;
• если k ( s ) = const > 0 и ?( s ) = const ? 0 , то ?( s ) – винтовая линия, см. рис. 4.
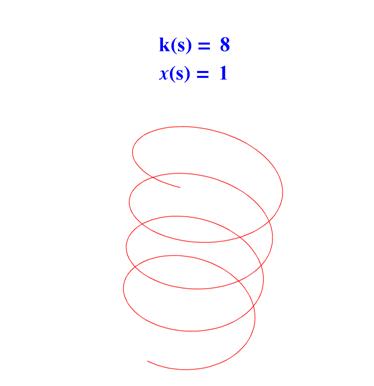
Рис. 4. Винтовая линия
На самом деле, соотношение между видом функций кривизны и кручения, с одной стороны, и формой кривой с другой, оказывается крайне нетривиальным: функциям достаточно простого вида могут соответствовать сложно устроенные кривые и обратно. Проиллюстрируем это, приведя еще несколько примеров. Начнем с плоских кривых.
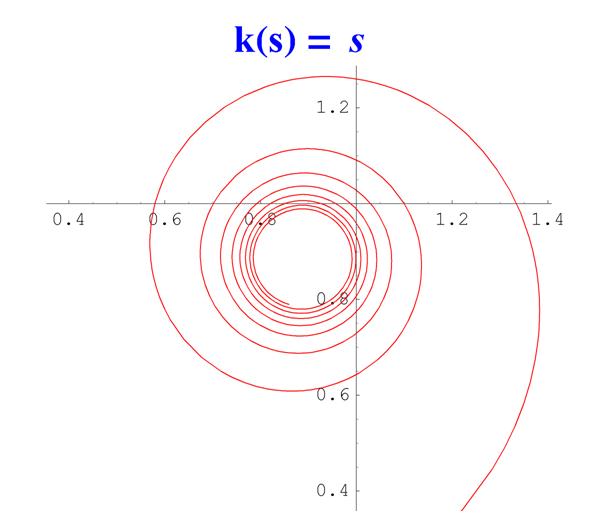
Рис. 5. Спираль
Отметим, что простым функциям кривизны часто соответствуют сложно устроенные кривые. Для иллюстрации приведем серию плоских кривых с кривизной вида k ( s ) = a cos 2 ( s ) + 1 .
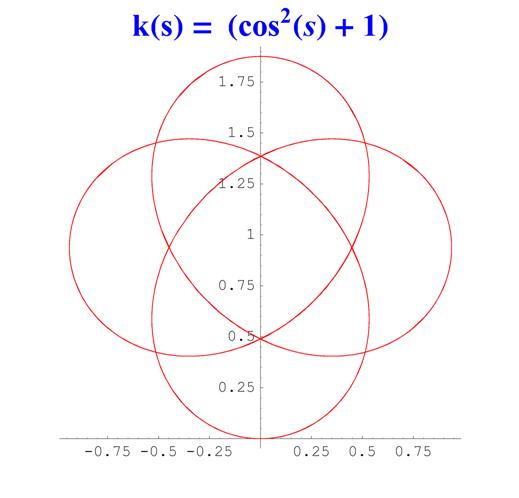
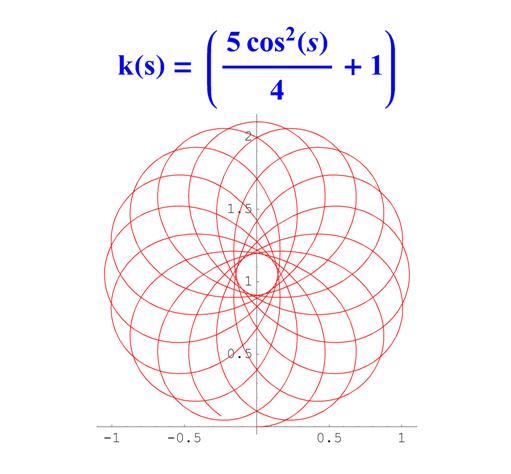
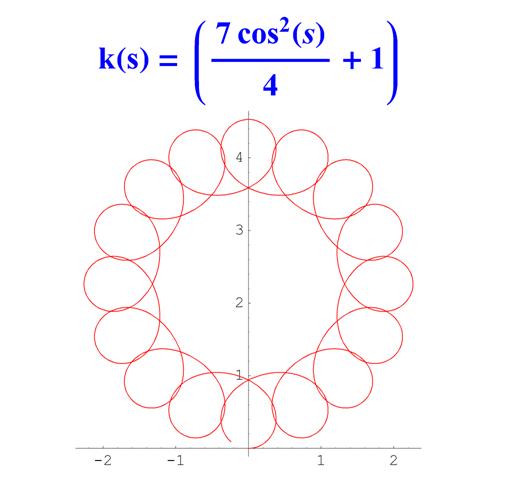
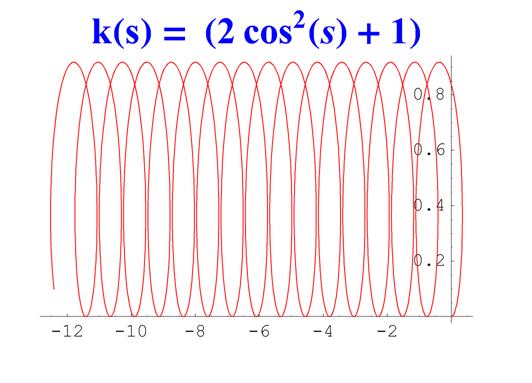
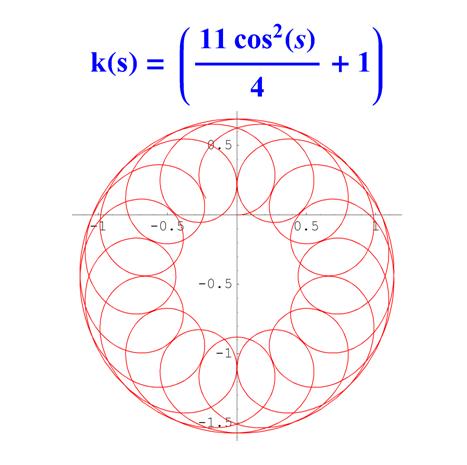
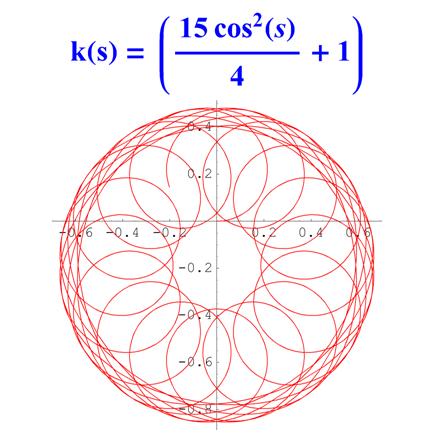
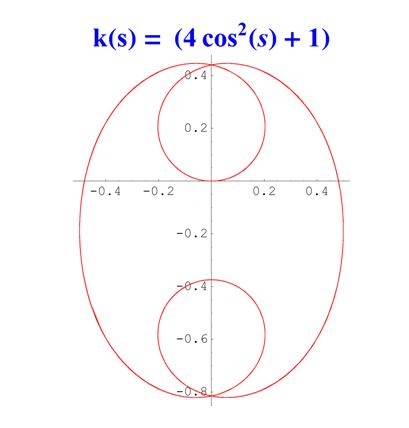
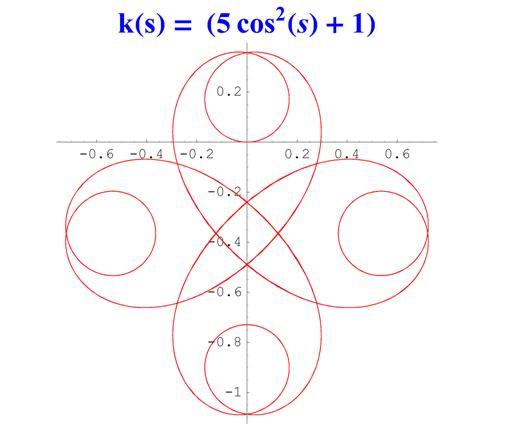
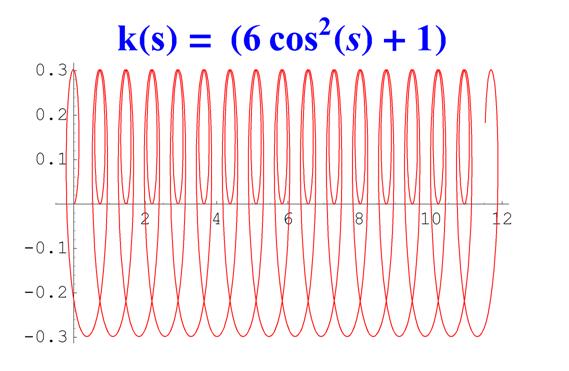
Рис. 6. Графики кривых
А вот пример кривой с чуть более сложно устроенной кривизной.
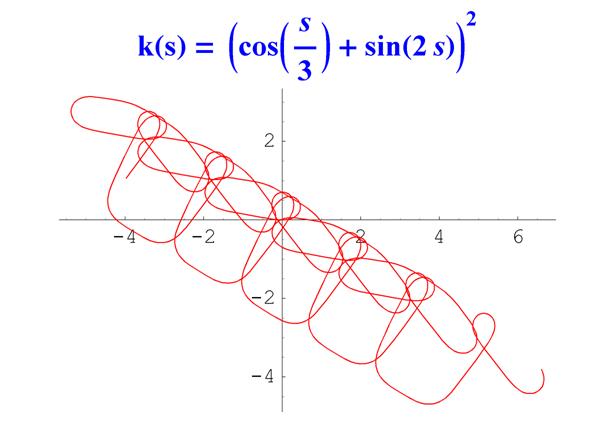
Рис. 7. График кривой
В случае пространственных кривых особенно наглядно сложность зависимости между кривизной, кручением и формой кривой можно увидеть на примере того, как деформируется кривая при изменении этих функций. Ниже приведена серия кадров анимации. Слева на каждом кадре изображена кривая, а справа – графики ее кривизны (красным цветом) и кручения (зеленым цветом). Обращаем внимание на то, что кривизны и кручения в данном случае задаются синусами и косинусами с меняющимися периодами, т.е. как сами функции, так и их изменения устроены достаточно просто. Тем не менее, соответствующие кривые выглядят крайне нетривиально (при просмотре анимационного ролика возникает впечатление, что кривая – это змея, извивающаяся под звуки флейты факира).
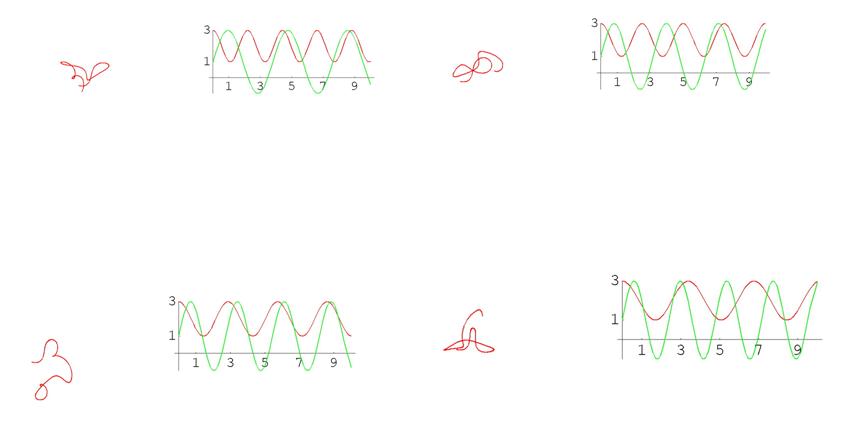
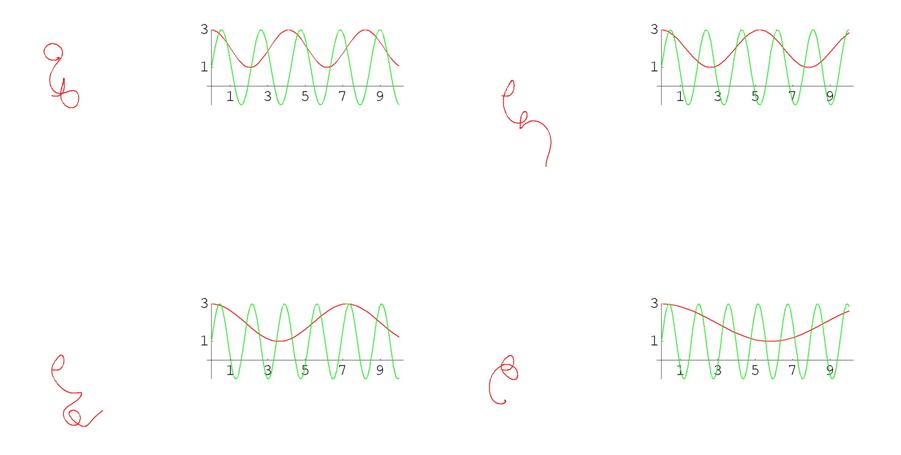
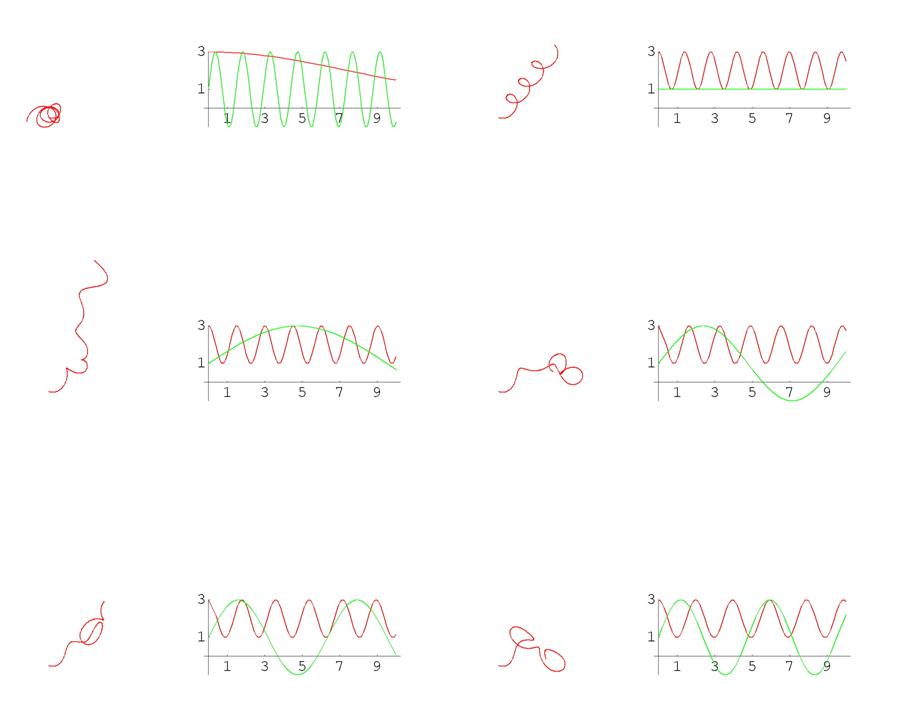
Рис. 8 Графики кривых
Еще один пример. Справа на графике красным цветом показана кривизна, а черным – кручение кривой, изображенной слева (кручение не меняется).
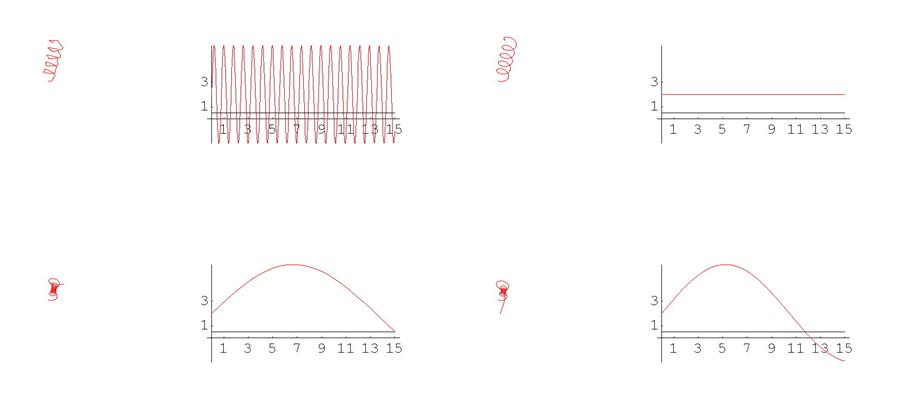
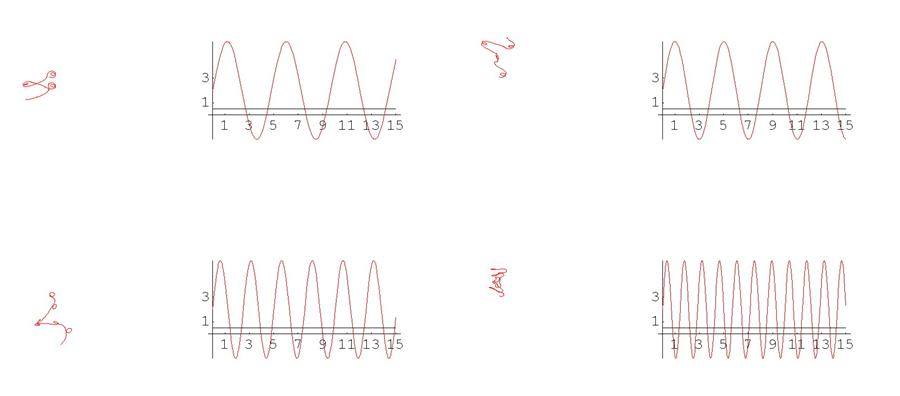
Рис. 9. Графики кривых
Таким образом, кривизна и кручение являются удобными характеристиками кривой, которые могут существенно упростить описание и исследование пространственной формы.
Вернемся теперь к изучению ломаных. Обсудим для начала, как можно определить характеристики ломаной, которые связаны с геометрией ломаных так же, как кривизна и кручение с геометрией кривых . Одним из критериев естественности определения является требование того, чтобы для каждой фиксированной бирегулярной кривой кривизна и кручение ломаной, вписанной в кривую, стремились бы к кривизне и кручению кривой измельчения ломаной. В действительности, существует много таких функций. Вот некоторые примеры:
• кривизна =
; кручение =
(несложно проверить, что после такой нормировки кривизна и кручение ломаной уже обладают требуемыми предельными свойствами);
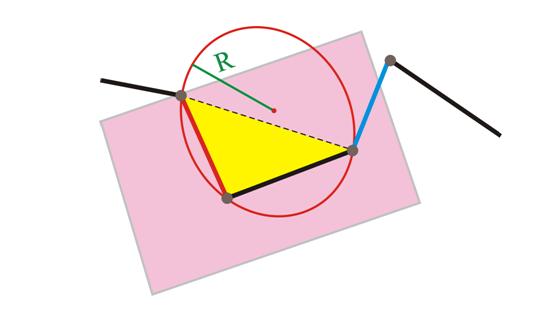
• определим кривизну во внутренней вершине как величину, обратно пропорциональную радиусу окружности, описанной вокруг треугольника, вершины которого – предыдущая, данная и последующая вершины ломаной;
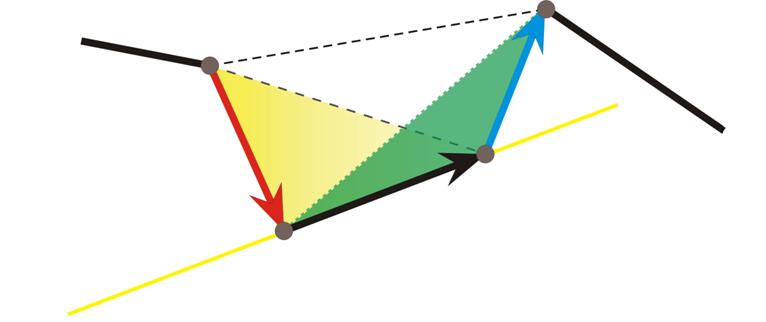
• кручение определим как величину, пропорциональную объему тетраэдра, натянутого на три последовательных звена ломаной. Объем дел ен им на произведение площадей граней этого тетраэдра, примыкающих к среднему звену;
• в случае равнозвенных ломаных первое определение можно использовать без деления на длины ребер, т.е. когда в качестве кривизны и кручения используются просто углы первого и второго типа. Получаются формулы, более простые для вычислений.
Ниже приводятся результаты численного эксперимента, демонстрирующего вид определенных выше аналогов кривизны и кручения для фрагментов реальных белков.
Рис. 10. Результаты численного эксперимента, демонстрирующего вид определенных выше
аналогов кривизны и кручения для фрагментов реальных белков.
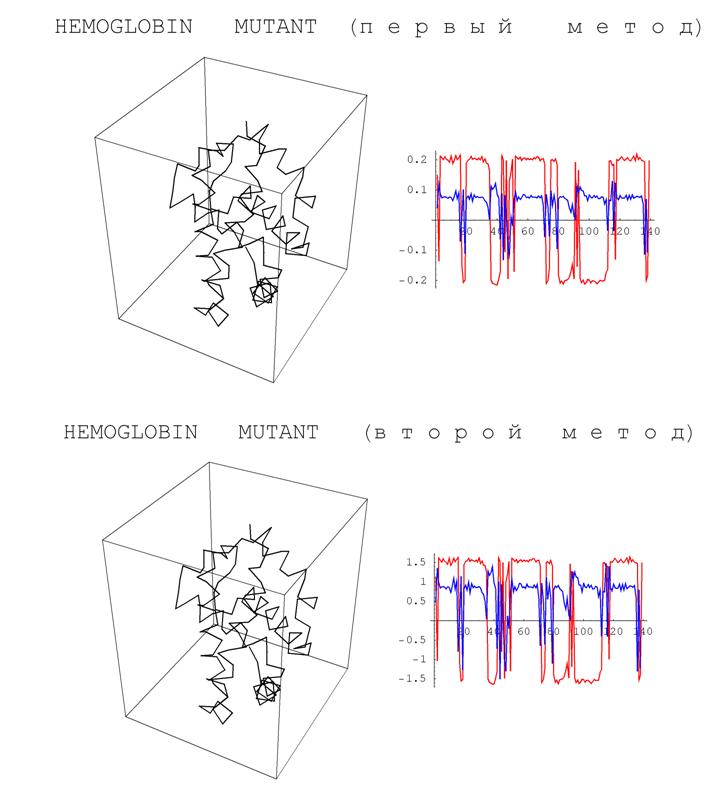
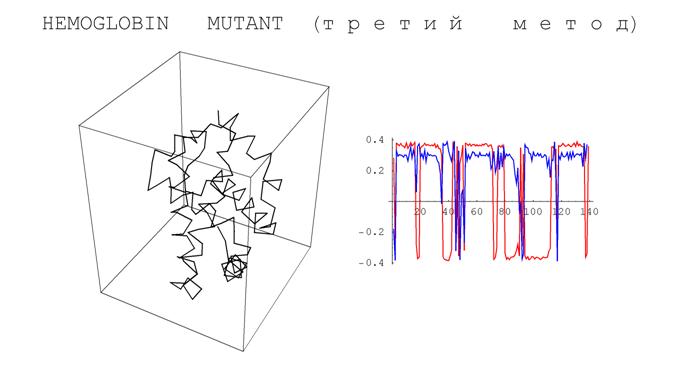
Заметим, что с точностью до скалярного множителя полученные графики похожи друг на друга. Впрочем, этот эффект объясняется отмеченным выше свойством моделирующих белки ломаных иметь звенья почти одинаковой длины.
Напомним, что постоянство кривизны и кручения регулярных кривых является характеристическим свойством винтовых линий. У белков эти винтовые линии возникают в виде a -спиралей. На полученных графиках участки, соответствующие a -спиралям отчетливо видны – им соответствуют почти горизонтальные фрагменты. Кроме того, плоские фрагменты кривых характеризуются обнулением кручения. В белках такие фрагменты называются b -слоями. На приведенных графиках b -слои выделяются фрагментами, на которых кручение (синяя кривая) осциллирует в окрестности нуля. Тем самым, одно из возможных приложений построенных характеристик – автоматическое выделение a -спиралей и b -слоев.
Еще одно возможное приложение – статистическое исследование связи параметров кривизны и кручений ломаной, описывающей белковую молекулу, с конкретными последовательностями аминокислот этой молекулы. Однако здесь требуется более сложная модель, разработкой которой занимается наша группа в настоящее время.
Также можно использовать построенные характеристики для моделирования изменения конформации белков. Идея состоит в том, чтобы устроить непрерывную деформацию кривизны и кручения начальной ломаной в кривизну и кручение конечной ломаной в подходящем классе функций, и для каждого набора промежуточных значений кривизны и кручения построить соответствующую ломаную. Ниже приводятся возможные реализации.
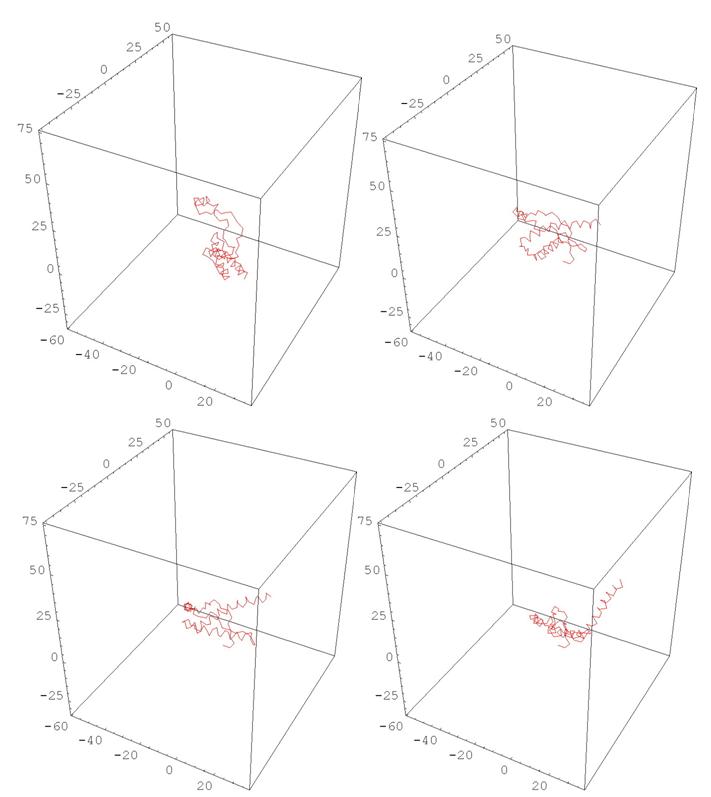
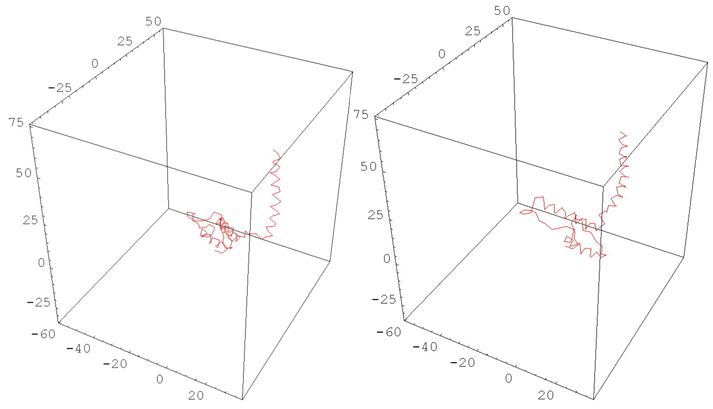
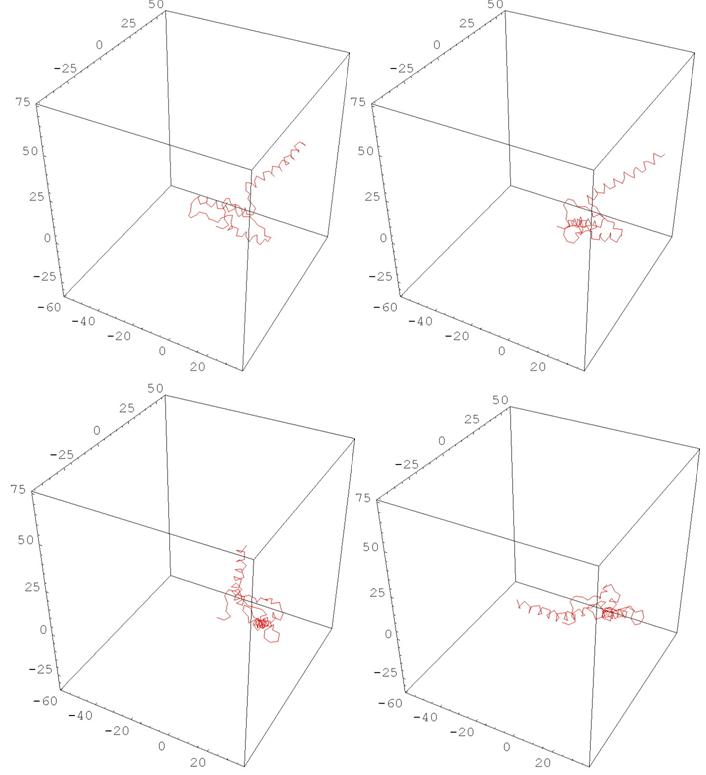
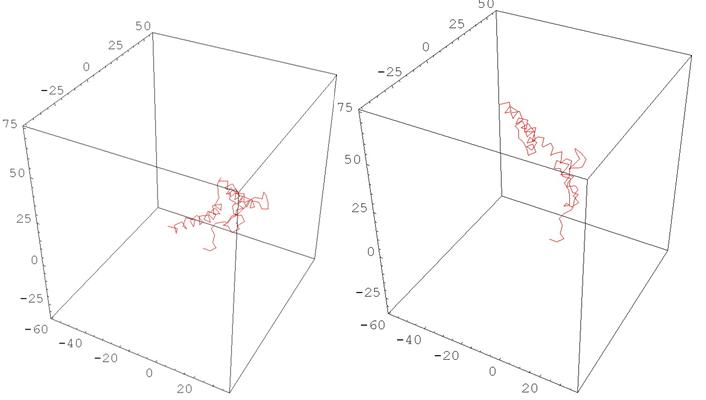
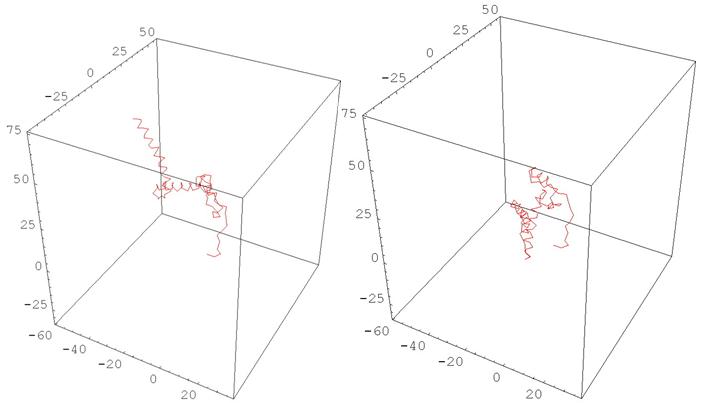
Рис. 11 HUMAN PRION PROTEIN
(деформация, меняющая одновременно все кривизны и кручения)
Еще одним примером использования геометрического подхода является построение механической модели пространственной структуры белка, которая основана на модели упругой фермы, составленной из узлов соединяющих их жестких стержней. Предполагается, что в узлах расположены атомы белка (альфа-углероды), а связи интерпретируются как состояние стержней, соединяющих узлы. Предполагается, что деформации фермы зависят от сил, приложенных к узлам, и подчиняются некоторому закону Гука. Силы, приложенные к узлам, порождаются силами притяжения или отталкивания некоторых атомов. Все связи делятся на валентные, пептидные, которые формируют структуру фермы, и водородные, слабые связи, которые деформируют стержни, а в конечном счете и всю ферму.
Расчет статического состояния упругой фермы приводит к нелинейным уравнениям, что приводит к методу линеаризации задачи и, тем самым, к расчету динамики формирования пространственной структуры молекулы. Отличие от существующих методов вычисления пространственной структуры белка заключается в том, что предлагаемая модель позволяет проводить вычисления при помощи быстрых алгоритмов, сложность которых оценивается порядком O(n 2 ) , где n – длина сиквенса пептида.
3. Формообразование тканей
Среди фундаментальных вопросов современной биологии одними из важнейших являются проблемы морфогенеза. Образование органов и тканей в процессе развития организма или при их регенерации демонстрирует поразительную воспроизводимость биологических структур с сохранением всех их основополагающих характеристик, включая клеточный состав, геометрические закономерности, соразмерность их субкомпонентов. Несмотря на интенсивное изучение этих процессов, история которого насчитывает уже более двухсот лет, до сих пор принципиальные моменты формообразования у биологических объектов остаются не раскрытыми. По-видимому, не существует единого механизма, и регуляция морфогенеза протекает на самых разных уровнях с участием различных физических и химических факторов. К ним можно отнести химические градиенты, рецепторное взаимодействие клеток и т.п. Таким образом, построение картины регуляции морфогенеза, а главное – принцип образования конкретной формы при этом процессе, наталкиваются на значительные трудности, связанные с многокомпонентностью и сложностью этой системы. В то же время, можно предположить, что на определенных этапах участвуют далеко не все факторы регуляции, или, по крайней мере, влиянием многих из них можно, с определенным приближением, пренебречь. Весь комплекс сложных механизмов регуляции направлен на реализацию относительно простых способов сборки трехмерной структуры, которые пока не раскрыты из-за сложностей всей системы. Это предположение открывает путь к построению относительно простых моделей, основанных на ограниченном наборе факторов регуляции.
Объект моделирования. Такие биологические структуры как органы и ткани обобщенно можно представить как сочетание двух компонентов: стромы органа и клеток различной дифференцировки. Мы не рассматривали влияния стромы и ограничились изучением клеточных ансамблей. Если задать вопрос, что же именно моделируется при таком подходе, то мы можем указать, по крайней мере, одно точное соответствие – многие из низших живых организмов, таких как, например, губка, способны восстанавливать свою структуру после разделения на отдельные клеточные элементы. Конечно, выделить такую ситуацию в более сложных многоклеточных организмах непросто, но и для них рассматриваемые механизмы могут иметь место. Кроме того, модели самосборки могут оказаться полезными и на другом уровне биологической организации – субклеточном. Образование многих надмолекулярных комплексов происходит именно путем самосборки из относительно простых и однородных компонентов.
Предлагаемая модель основана на гипотезе о том, что форма ткани однозначно задается структурой распределения на поверхности клеток точек межклеточных контактов (например, плотных контактов). Плотные контакты являются одним из главных компонентов межклеточного взаимодействия и выполняют ряд важных функций. Наличие таких межклеточных контактов обязательно для большинства типов тканей. Тем самым, задача сводится к исследованию связи между различными вариантами пространственного расположения точек крепления (например, плотных контактов на поверхности клетки) и формирующимися пространственными многоклеточными структурами-формами.
В рамках этой гипотезы нами была написана программа, позволяющая моделировать различные ткани в зависимости от структуры точек крепления. Простейшая версия нашей модели такова.
Клетки моделируются сферами некоторого радиуса (в простейшем случае все радиусы одинаковы). Структура точек крепления задается конечным набором точек на единичной сфере (модельной сфере). В начальный момент времени мы задаем случайные положения некоторого количества клеток координатами центров соответствующих сфер. Кроме того, для каждой такой клетки мы задаем случайно ориентированную структуру точек крепления, выполняя отображение подобия с модельной сферы на сферу, задающую клетку. При этом имеется возможность задавать клетки нескольких разных типов, т.е. с разными структурами точек крепления.

Рис. 12. Рабочий экран программы, моделирующей тканеобразование. Ткань из клеток 3-х типов.
Затем запускается модуль, имитирующий деление клеток для формирования ткани. Программа перебирает имеющиеся клетки и для каждой из них просматривает свободные точки крепления. Если такая точка найдена, то в систему добавляется новая клетка, которая касается данной по выбранной точке крепления, при условии, что эта новая клетка не пересекает внутренностей уже построенных клеток. Если новая клетка добавлена, структура ее точек крепления может быть устроена по-разному. В простейшей ситуации точки крепления порождаются симметрией относительно общей касательной плоскости в точке касания исходной и построенной клеток. Также имеется возможность задать случайный поворот вокруг линии центров. Кроме того, предусмотрен специальный режим, учитывающий, что деление данной клетки возможно лишь при некотором ее специальном пространственном положении, например, радиус-вектор некоторого рецептора составляет с выделенным направлением острый угол.
Мы описали лишь ключевые идеи, заложенные в программу. На самом деле, имеется много разных параметров, которые управляют процессом формообразования. Так, например, чтобы отразить в модели неточечное касание клеток, форма которых, может, отличается от сферической, мы допускаем пересечение клеток-сфер с заранее заданным ограничение на глубину. Это делает картину более реалистичной.
Преимущество программы состоит в том, что в процессе формообразования разрешается менять структуру точек касания, что мгновенно модифицирует изображенную ткань и, тем самым, позволяет эффективно увидеть соответствие между локальной геометрией точек крепления и глобальными свойствами соответствующей ткани.
При этом удается получать как регулярные пространственные структуры, так и хаотичные. Ниже приведен ряд примеров полученных нами конфигураций.
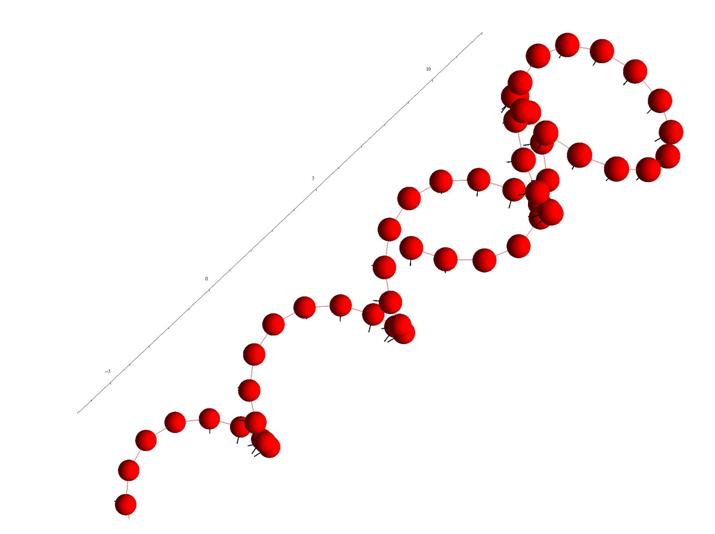
Рис. 13. Спиралеобразная ткань из клеток одного типа.
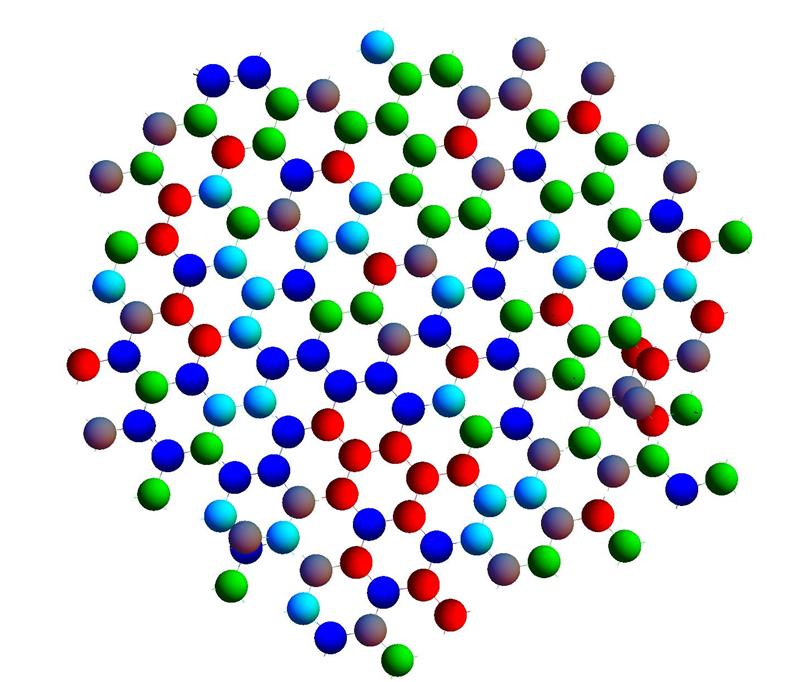
Рис. 14. Почти плоская ткань из клеток нескольких почти одинаковых типов.
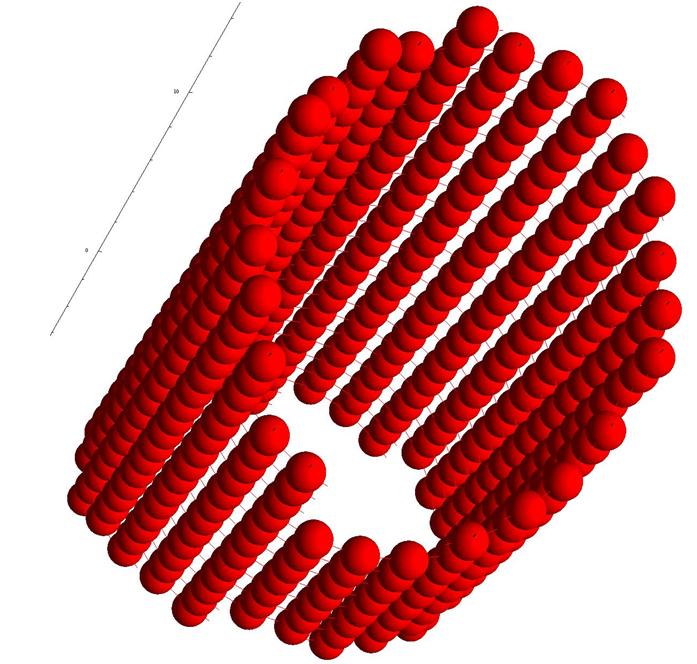
Рис.15. Цилиндр из клеток одного типа.

Рис.16. Скрученная двойная спираль.
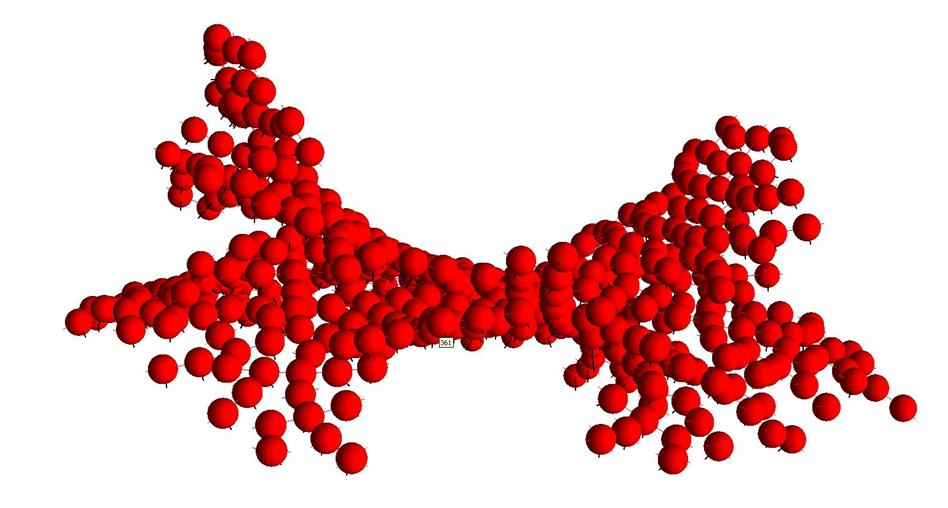
Рис.17. Седловидная поверхность из клеток одного типа. Номер указывает на клетку, полученную 361-ой.
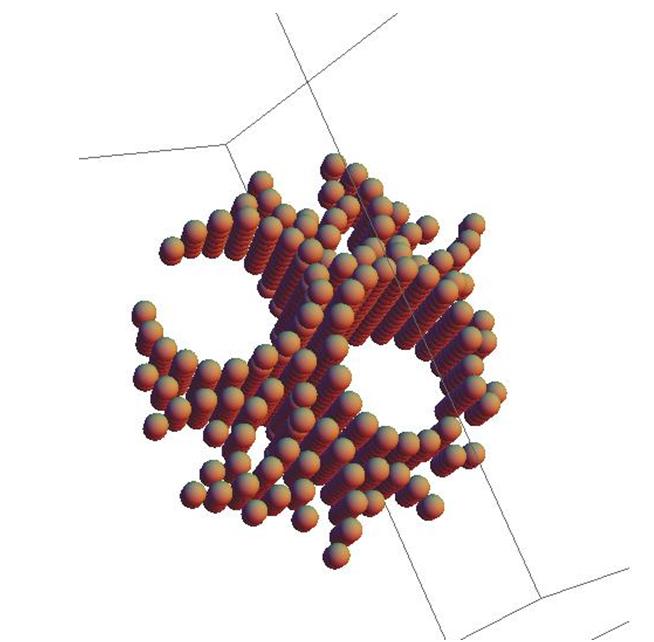
Рис.18. Двойной цилиндр.
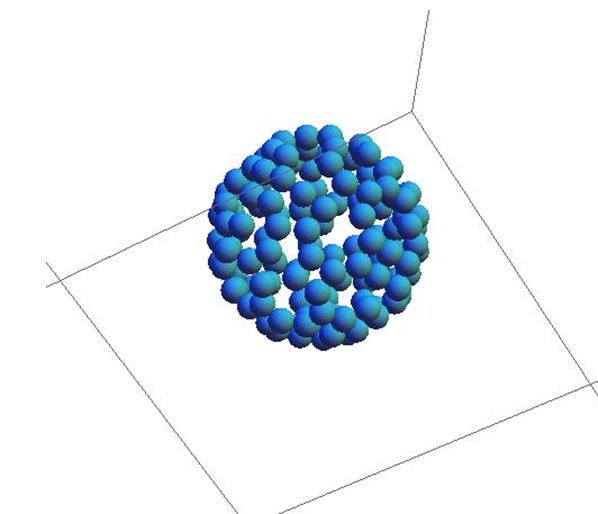
Рис. 19. Сфера, составленная из клеток одного типа.
Таким образом, мы видим, что использование простейших предположений о зависимости формирования биологических структур от пространственного расположения точек крепления на поверхности отдельных элементов, позволяет получать достаточно сложные образования.
Работа выполнена при частичной поддержке РФФИ (проекты № 07-01-00648, № 05-01-22002 НЦНИ), гранта Президента РФ поддержки ведущих научных школ РФ (проект НШ-660.2008.1), программы Эйлера ДААД, а также проекта Министерства науки и образования РФ РНП 2.1.1.7988.
Список литературы
1. А.М.Леонтович, И.И.Пятецкий-Шапиро, О.Н.Ставская, Некоторые математические задачи, связанные с формообразованием, Автоматика и телемеханика , 4 (1970), стр . 94-107.
2. Fold fully forecast. Computer predicts protein's shape from sequence alone. Nature News ( 23 Sep 2002 ), doi: 10.1038/news020923-4, News http://www.nature.com/news/2002/020923/full/news020923-4.html
3. К.В.Шайтан. Молекулярная динамика пептидов. http://www.moldyn.org/library/moldyn/moldyn.htm
4. К.В.Шайтан, С.С.Сарайкин, Метод молекулярной динамики, 1999, http://www.moldyn.ru/library/md/default.htm
5. А.В.Финкельштейн. Введение в физику белка. Курс лекций 1999-2000.
6. А.В.Финкельштейн, О.Б.Птицын ``Физика белка'', Москва, ``Книжный дом ``Университет'''', 2002.